Anti-FDX1 - Ferredoxin

Cat# AS06121
Size : 50ul
Brand : Agrisera
Anti-FDX1 | Ferredoxin
AS06 121 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. thaliana, H. vulgare, P. strobus, S. oleracea, Ch. reinhardtii, Synechocystis 6803 substrain PCC-M
-
This product doesn't have any reviews.
| Immunogen: | Native ferredoxin purified from Spinacia oleracea. |
| Host: | Rabbit |
| Clonality: | Polyclonal |
| Purity: | Serum |
| Format: | Lyophilized |
| Quantity: | 50 µl |
| Reconstitution: | For reconstitution add 50 µl of sterile water |
| Storage: | Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. |
| Tested applications: | Western blot (WB) |
| Recommended dilution: | 1 : 1000 (WB) |
| Expected | apparent MW: | 10 (Spinacia oleracea); 16.7 (Arabidopsis thaliana), 13.7 (Chlamydomonas reinhardtii) |
| Confirmed reactivity: | Arabidopsis thaliana, Chlamydomonas reinhardtii, Hordeum vulgare, Pinus strobus, Spinacia oleracea, Synechocystis 6803 substrain PCC-M |
| Predicted reactivity: | Dicots, Nicotiana tabacum, Physcomitrium patens, Zea mays Species of your interest not listed? Contact us |
| Not reactive in: | Phaeodactylum tricornutum |
|
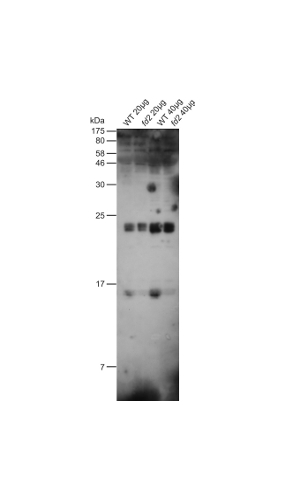
| Additional information: | In Arabidopsis thaliana leaf extracts there is a strong cross-reactivity at 20 kDa. |
| Additional information (application): | Load per well: 20-40 µg/lane required for Arabidopsis thaliana, 2-10 µg for other species. |
| Background: | Ferredoxins are acidic, low molecular weight, soluble iron-sulfur proteins found in various organisms. Iron-sulfur proteins are defined as proteins carrying iron-sulfur cluster(s) in which the iron is at least partially coordinated by sulfur. The protein acts as multifunctional electron carriers in diverse redox systems. The chloroplast ferredoxin is involved in both cyclic and non-cyclic photophosphorylation reactions of photosynthesis and other reductive reactions in the chloroplast. |
| Selected references: | Tiwari et al. (2024).Differential FeS cluster photodamage plays a critical role in regulating excess electron flow through photosystem I. Nat Plants. 2024 Oct;10(10):1592-1603. doi: 10.1038/s41477-024-01780-2. Lin et al. (2024). Hydrogen production in the Chlorella sp. DT mutants carrying heterologous electron donor ferredoxin 1 of Chlamydomonas reinhardtii. Electronic Journal of Biotechnology Volume 69, May 2024, Pages 11-20. Cvetkovska et al. (2018). Characterization of photosynthetic ferredoxin from the Antarctic alga Chlamydomonas sp. UWO241 reveals novel features of cold adaptation. New Phytol. 2018 Jul;219(2):588-604. doi: 10.1111/nph.15194. Jokel et al. (2018). Hunting the main player enabling Chlamydomonas reinhardtii growth under fluctuating light. Plant J. 2018 Mar 25. doi: 10.1111/tpj.13897. Georg et al. (2017). Acclimation of Oxygenic Photosynthesis to Iron Starvation Is Controlled by the sRNA IsaR1. Curr Biol. 2017 May 22;27(10):1425-1436.e7. doi: 10.1016/j.cub.2017.04.010. Hu et al. (2017). The SUFBC2 D Complex is Required for the Biogenesis of All Major Classes of Plastid Fe-S Proteins. Plant J. 2017 Jan 19. doi: 10.1111/tpj.13483. Higuchi et al. (2011). Modulation of macronutrient metabolism in barley leaves under iron-deficient condition. Soil Sc and Plant Nutr. 57 (2): 233-247. |
Protocols to work with plant and algal protein extracts